Kompleks Golgi: opis
Kompleks Golgi je bil odkrit leta 1898. Ta membranska struktura je zasnovana tako, da izloča spojine, ki se sintetizirajo v endoplazmatskem retikulumu. Nadalje se bomo seznanili s tem sistemom. 
Kompleks Golgi: stavba
Naprava je kup membranskih rezervoarjev v obliki diska. Te vrečke so nekoliko razširjene proti robovom. Sistem mehurčkov Golgi je povezan s tanki. V živalskih celicah je prisoten en velik ali več pilotov, ki so povezani s cevmi, v rastlinske celice Dictyosomes so odkriti (več ločenih nizov). Kompleks Golgi vključuje tri oddelke. Obdani so z membranskimi mehurčki:
- cis-najbližje jedru;
- medial;
- oddelek (najbolj oddaljen od jedra).
Te sisteme odlikuje komplet encimov. V cis oddelku se prva vrečka imenuje "reševalna cisterna". S svojo pomočjo se receptorji, ki prihajajo iz endoplazmatskega vmesnega omrežja, premaknejo nazaj. Oddelek za encime cis, imenovan fosfoglikozidaza. Doda fosfat manozi (ogljikovim hidratom). V medialnem delu sta dva encima. To še posebej velja za mennadiazo in N-acetilglukozamin transferazo. Slednji dodaja glikozamine. Encimi trans-oddelka: peptidaza (izvaja proteolizo) in transferazo (s svojo pomočjo poteka prenos kemičnih skupin). 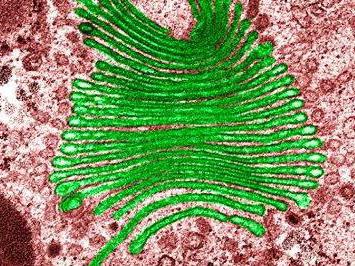
Golgijev kompleks: funkcije
Ta struktura zagotavlja ločevanje beljakovin v naslednje tri tokove:
- Lizosomal. V skladu s tem glikirani proteini prodrejo v cis-oddelek Golgijevega aparata. Nekatere izmed njih so fosforilirane. Posledično nastane manoza-6-fosfat - tržozosomski encimi. V prihodnosti bodo ti fosfatni proteini vstopili v lizosome in se ne bodo spreminjali.
- Konstitutivna eksocitoza (izločanje). Ta tok vključuje beljakovine in lipide, ki so postali sestavni deli aparata celične površine, vključno z glikokaliksom. Prav tako so lahko prisotne spojine, ki so del zunajceličnega matriksa.
- Inducirano izločanje. Proteini, ki delujejo zunaj celice, površinski aparat, prodrejo v ta tok v notranjem okolju telesa. Inducirana sekrecija je značilna za sekrecijske celice.
Kompleks Golgi sodeluje pri tvorbi sluznice - mukopolisaharidi (glikozaminoglikani). Naprava oblikuje tudi ogljikohidratne sestavine glikokaliksa. Večinoma so to glikolipidi. Sistem zagotavlja tudi sulfatacijo beljakovinskih in ogljikohidratnih elementov. Kompleks Golgi sodeluje pri delni proteolizi beljakovin. V nekaterih primerih zaradi tega spojina iz neaktivnega preide v aktivno obliko (npr. Proinzulin se pretvori v insulin). 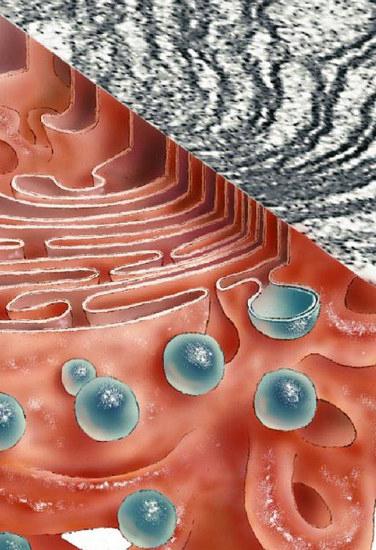
Premikanje spojin iz endoplazmatskega retikuluma (EPS)
Kompleks je asimetričen. Bližje jedru celice so najbolj nezrele beljakovine. Vezikli, membranski vezikli, so nenehno pritrjeni na te vrečke. Odplazijo iz endoplazmatskega granularnega retikuluma. Na njenih membranah poteka proces sinteze beljakovin z ribosomi. Transport spojin iz endoplazmatskega retikuluma v kompleks Golgi je nediskriminatoren. Hkrati pa še naprej ostajajo nepravilno ali nepopolno zložene beljakovine v EPS. Obratno gibanje spojin v endoplazmatski retikulum zahteva posebno signalno sekvenco in je omogočeno z vezavo teh snovi na membranske receptorje v cis-predelu.
Spreminjanje beljakovin
V rezervoarjih kompleksnega zorenja nastanejo spojine, ki so namenjene za izločanje, transmembransko, lizosomsko in druge snovi. Ti proteini se zaporedno premikajo vzdolž tankov na organele. Njihove spremembe se začnejo v njih - fosfoliranje in glikozilacija. Med prvim postopkom se na proteine pritrdi ostanek. fosforna kislina. V O-glikozilaciji so kompleksni sladkorji zasidrani preko kisikovega atoma. Različni rezervoarji vsebujejo različne katalitske encime. Posledično potekajo postopno različni procesi z zorenjem beljakovin v njih. Nedvomno je treba nadzorovati takšen stopničast pojav. Kot vrsta "oznake kakovosti" se uporabljajo polisaharidni ostanki (predvsem manoza). Označujejo zrelosti beljakovin. Nadaljnje gibanje skozi rezervoarje spojin znanost v celoti ne razume, kljub dejstvu, da so odporne snovi manj ali bolj povezane z eno vrečko. 
Prevoz proteinov iz aparata
Iz zapletenih mehurčkov za presadke. Vsebujejo popolnoma zrele proteinske spojine. Glavna naloga kompleksa je razvrščanje beljakovin, ki prehajajo skozi njega. V aparatu nastane "tri smerni proteinski tok" - zorenje in transport:
- Spojine plazemske membrane.
- Skrivnosti.
- Lizosomski encimi.
Skozi vezikularni transport se proteini, ki so prešli skozi kompleks Golgi, dostavljajo na določena mesta v skladu z "oznakami". Ta proces prav tako ni popolnoma razumljen. Ugotovljeno je bilo, da je za prenos beljakovin iz kompleksa potrebno sodelovanje specifičnih membranskih receptorjev. Prepoznajo spojino in zagotavljajo selektivno spajanje vezikla in enega ali drugega organela. 
Nastanek lizosomov
Skozi aparat prehaja veliko hidroliznih encimov. Dodajanje zgoraj navedene oznake poteka ob sodelovanju dveh encimov. Specifično prepoznavanje lizosomskih hidrolaz z elementi njihove terciarne strukture in dodajanje N-acetilglukozamin fosfata se izvaja z N-acetilglukozamin fosfotransferazo. Fosfoglikozid, drugi encim, odstrani N-acetilglukozamin, kar povzroči nastanek oznake M6F. Po drugi strani ga prepoznajo receptorski proteini. S svojo pomočjo se hidrolaze prenašajo v mehurčke in njihove mandate v lizosome. V kislih pogojih se fosfat odcepi od zrele hidrolaze v njih. Če obstajajo motnje v aktivnosti N-acetilglukozamin fosfotransferaze zaradi mutacij ali zaradi genetskih okvar v M6F receptorju, se vsi lizosomski encimi privzeto oddajo zunanji membrani. Nato se izločajo v zunajcelične razmere. Prav tako je bilo ugotovljeno, da se nekateri M6P receptorji prenašajo tudi na zunanjo membrano. Med endocitozo izvedejo vrnitev naključno ujetih lizosomskih encimov iz zunanjega okolja v celici. 
Prevoz snovi do zunanje membrane
Običajno so tudi v fazi sinteze v steno endoplazmatskega retikuluma vgrajene proteinske spojine zunanje membrane z njihovimi hidrofobnimi regijami. Nato se dostavijo v kompleks Golgi. Od tam se prevažajo na celično površino. V procesu fuzije plazmaleme in veziklov se takšne spojine ne sproščajo v okolje.
Izločanje
Skoraj vse spojine, proizvedene v celici (tako beljakovinske kot ne-proteinske narave), preidejo skozi kompleks Golgi. Tam se razvijejo v sekretorne vezikule. Pri rastlinah z diktijami se tako proizvaja material. celična stena.