Dvojno gnojenje v rastlinah
Dvojno gnojenje v rastlinah je velikega biološkega pomena. Navašin ga je odkril leta 1898. Nato podrobneje pogledamo, kako se v rastlinah pojavlja dvojna gnojenje. 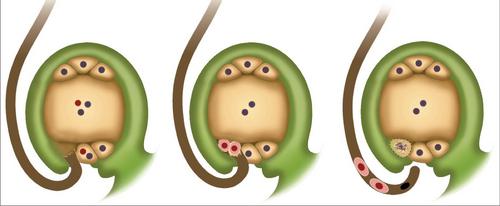
Biološki pomen
Proces dvojne gnojenja spodbuja aktivni razvoj hranilnega tkiva. V tem smislu jajčec ne shranjuje materiala za prihodnost. To pa pojasnjuje njegov hiter razvoj.
Shema dvojnega gnojenja
Na kratko, pojav lahko opišemo na naslednji način. Dvojno oploditev kritosemenk se sestoji iz prodiranja dveh spermijev v jajčnik. Ena se združi z jajcem. To prispeva k začetku razvoja diploidnega zarodka. Druga sperma se poveže z osrednjo celico. Posledično se tvori triploidni element. Iz te celice se pojavi endosperm. Je hranilni material za razvijajoči se zarodek.
Razvoj cvetnega prahu
Dvojna oploditev kritosemenk se začne po nastanku močno zmanjšane haploidne generacije. Predstavljajo ga gametofiti. Dvojno gnojenje cvetočih rastlin spodbuja kalitev peloda. Začne se z nabrekanjem zrn in posledično nastajanjem cvetnega prahu. V najtanjšem delu se prebije skozi sporoderm. Imenuje se zaslonka. S konice cvetnega prahu se oddajajo posebne snovi. Zmehčajo tkiva kolone in stigmatizirajo. Zaradi tega vključujejo cvetni prah. Ko se razvija in raste, v njej prehajajo tako sperma kot jedro iz vegetativne celice. V veliki večini primerov cvetni prah prodre v nucelus (megasporangia) skozi mikropil jajc. To se redko dogaja na drug način. Po prodiranju v zarodno vrečko se cepi cvetni prah. Posledično se vsa njena vsebina vlije v notranjost. Dvojna oploditev cvetočih rastlin se nadaljuje z nastankom diploidnega zigota. To prispeva k prvi spermi. Drugi element je povezan s sekundarnim jedrom, ki se nahaja v osrednjem delu zarodka. Nastalo triploidno jedro se nato transformira v endosperm. 
Oblikovanje celic: splošne informacije
Proces dvojne oploditve cvetočih rastlin se izvaja s posebnimi zarodnimi celicami. Njihova tvorba poteka v dveh fazah. Prva faza se imenuje sporogeneza, druga je hematogeneza. V primeru nastanka moških celic se te faze imenujejo mikrosporogeneza in mikroematogeneza. Pri oblikovanju ženskih genitalnih elementov se predpona spremeni v "mega" (ali "makro"). Sporogeneza temelji na mejozi. To je proces nastajanja haploidnih elementov. Pred mejozo, pa tudi med predstavniki favne, sledi razmnoževanje celic skozi mitotične delitve.
Proizvodnja semenčic
Primarno nastajanje moških genitalnih elementov se izvaja v posebnem bradavičnem tkivu. Imenuje se archesparial. V njem zaradi mitoze nastajajo številni elementi, matične celice peloda. Nato vstopijo v mejozo. Zaradi dveh meiotičnih delitev nastanejo 4 haploidne mikrospore. Nekaj časa ležijo v bližini in tvorijo tetrado. Po tem se razgradijo v zrnca cvetnega prahu - posamezne mikrospore. Vsak od oblikovanih elementov se začne prekrivati z dvema lupinama: zunanjim (exine) in notranjim (intin). Nato se začne naslednja faza - mikrogametogeneza. Ta pa je sestavljena iz dveh mitotičnih zaporednih delitev. Po nastanku prvih dveh celic: generativne in vegetativne. Nato prvi preide v drugo delitev. Tako nastanejo dve moški celici - semenčice. 
Makroporogeneza in megasporogeneza
V tkivih ovul se začne ločevati eden ali več arksporijskih elementov. Hitro rastejo. Zaradi te aktivnosti postanejo bistveno večje od preostalih celic, ki jih obdajajo v jajcih. Vsak element arpspore podvržemo delitvi z mitozo enkrat, dva ali večkrat. V nekaterih primerih lahko celica takoj preide v mater. Mejoza se pojavi znotraj nje. Tako nastanejo 4 haploidne celice. Praviloma se največji od njih začne razvijati in se spreminja v zarodno vrečko. Trije ostajajo postopoma degenerirani. Na tej stopnji je makrosporogeneza zaključena, začne se makro hematogeneza. V njem se pojavijo mitotične delitve (večina kritosjemcev ima tri). Citokineza ne spremlja mitoze. Zaradi treh delitev nastane zarodna vrečka z osmimi jedri. Kasneje se izolirajo v neodvisne celice. Ti elementi so na določen način porazdeljeni preko zarodne vrečke. Ena od izoliranih celic, ki je pravzaprav jajčna celica, skupaj z dvema drugim sinergidoma, poteka v mikropilu, v katerega pride do vdora sperme. V tem procesu imajo sinergisti zelo pomembno vlogo. Vsebujejo encime, ki prispevajo k raztapljanju membran na cvetni prah. Na nasprotni strani zametka se nahajajo druge tri celice. Imenujejo se antipodi. S pomočjo teh elementov se hranila prenesejo iz ovul v zarodno vrečko. Preostale dve celici se nahajata v osrednjem delu. Pogosto se združijo. Zaradi njihove povezave nastane diploidna centralna celica. Ko pride do dvojne oploditve in sperme prodrejo v jajčnik, se ena od njih, kot je omenjeno zgoraj, združi z jajčno celico. 
Značilnosti cvetnega prahu
Dvojno gnojenje spremlja njegova interakcija s sporofitnimi tkivi. To je precej specifično. Ta proces ureja delovanje kemičnih spojin. Ugotovljeno je, da če je cvetni prah noter destilirana voda izgubila bo sposobnost kalivosti. Če je nastala raztopina koncentrirana in nato predelana, bo ponovno postala popolna. Razvoj cvetnega prahu po kalivosti nadzoruje tkivo pestiča. Na primer, v bombažu, njegova rast za jajce traja približno 12-18 ur. Vendar pa je že po 6 urah možno ugotoviti, na katero jajce bo šla cvetni prah. To je razumljivo, ker se v njem začne uničevanje sinergije. Trenutno ni ugotovljeno, kako lahko obrat usmerja razvoj cevi v pravo smer in kako se sinergist nauči o približevanju. 
"Prepoved" samopodobe
Pogosto ga opazimo v cvetočih rastlinah. Ta pojav ima svoje značilnosti. »Prepoved« za samoopraševanje se kaže v dejstvu, da sporofit »identificira« svojega moškega hematofita in mu ne dovoljuje sodelovanja pri oploditvi. Istočasno se v nekaterih primerih ne pojavlja kalitev lastnega cvetnega prahu na stigmi pestiča. Vendar se praviloma rast cevi še vedno začne, vendar se kasneje ustavi. Posledica tega je, da cvetni prah ne doseže jajca in posledično ne pride do dvojne oploditve. Darwin je opazil ta pojav. Torej je našel v jaglaca spomladansko cvetje dve obliki. Nekateri so bili dolgi stebli s kratkimi prašniki. Druge so kratke. V njih so bili staminatni filamenti dolgi. Rastline s kratkimi stebli se razlikujejo po velikem cvetnem prahu (dvakrat toliko kot pri drugih). Hkrati so celice v papilah stigme majhne. Ti znaki so pod nadzorom skupine tesno tkanih genov. 
Receptorji
Dvojno gnojenje je učinkovito, kadar se cvetni prah prenaša iz ene oblike v drugo. Za prepoznavanje lastnih elementov so odgovorne posebne receptorske molekule. So kompleksne spojine ogljikovih hidratov z beljakovinami. Ugotovljeno je bilo, da se oblike divjega zelja, ki ne proizvajajo teh receptorskih molekul v tkivih stigme, lahko samoprašijo. Za normalne rastline se ogljikove hidratne proteinske spojine pojavijo dan pred odprtjem rože. Če dvakrat pred cvetenjem odprete brst in ga obdelate s svojim cvetnim prahom, se bo zgodilo dvojno gnojenje. Če se to stori dan pred otvoritvijo, ne bo.
Alleles
Omeniti velja, da je v nekaterih primerih "samo-nezdružljivost" cvetnega prahu v rastlinah določena z vrsto večkratnih elementov posameznega gena. Ta pojav je podoben nezdružljivosti tkivnih presaditev pri živalih. Takšni aleli so označeni s črko S. Število v teh populacijah lahko doseže desetine ali celo stotine. Na primer, če je genotip obrata za proizvodnjo jajc s1s2, in s2s3, ki proizvaja cvetni prah, med navzkrižnim opraševanjem, se kalivost opazi le v 50% prašnih delcev. To bodo tisti, ki nosijo alel s3. Če obstaja več ducatov elementov, bo večina cvetnega prahu normalno kalila med navzkrižnim opraševanjem, medtem ko je samoprašenje popolnoma preprečeno. 
Za zaključek
Za razliko od gnoserspermov, za katere je značilen razvoj dovolj močnega haploidnega endosperma, ne glede na oploditev, pri kritosemenkah se tkivo oblikuje samo v tem primeru. Glede na veliko število generacij je tako dosegel znatne prihranke energije. Povečanje stopnje ploidnosti endosperma očitno prispeva k hitrejši rasti tkiva v primerjavi z diploidnimi plasti sporofita.